Cell Research | 蓝斐团队与合作者揭示精子发生的表观遗传调控机制
精子发生是哺乳动物雄性生殖细胞发育分化的核心过程,涉及有丝分裂、减数分裂和精子形成三大最基本的细胞生物学事件。在有丝分裂阶段,精原细胞通过自我更新、增殖和分化,维持精子发生的连续性和稳定性;减数分裂阶段涉及同源染色体配对、重组交换和分离,确保遗传稳定性并促进遗传多样性;而精子形成阶段则伴随顶体形成、组蛋白-过渡蛋白-鱼精蛋白转换、核浓缩及染色质重塑等一系列复杂的细胞生物学和形态学变化,赋予精子运动和受精能力。
精子发生的精确调控依赖于基因表达的时空协调。然而,由于睾丸组织内同时存在多种不同发育阶段的生精细胞,且缺少相应的特异性标志物,高纯度、特定发育阶段生精细胞的分离长期以来一直面临技术挑战。因此,尽管已有研究揭示了部分发育阶段的基因调控机制,但对于精子发生全过程的基因表达调控仍缺乏系统性理解。
表观遗传修饰通过动态调控基因的激活与抑制,被认为在精子发生过程中发挥重要调控作用。然而,受制于前述原因,该过程中表观遗传修饰谱的动态变化及其功能机制尚未被系统地研究过。
2025年3月3日,复旦大学生物医学研究院蓝斐团队与中国科学院分子细胞科学卓越创新中心童明汉团队和北京大学汤富酬团队合作在Cell Research在线发表了题为SETD1B-Mediated Broad H3K4me3 Controls Proper Temporal Patterns of Gene Expression Critical for Spermatid Development 的研究成果。该研究首次绘制了覆盖小鼠精子发生全过程的表观遗传动态变化图谱,系统揭示了表观遗传修饰在精子发生过程中的动态变化过程和规律;亮点发现为:1. 发现了一种特殊存在于圆形精子细胞阶段的broad H3K4me3修饰模式(组蛋白H3第4位赖氨酸三甲基化的宽泛修饰区域,大多长度介于5-10kb),该修饰模式由RFX2-SETD1B轴介导,发生在数千个增强子和启动子区,通过与常见的1-2kb长度的典型H3K4me3(typical H3K4me3)竞争转录机器,负责调控精子形成关键基因的高效表达和精准时序调控;2. 发现在有丝分裂向减数分裂转换时以及减数分裂重组完成后,H3K27me3和H3K9me2的出现时间呈现显著互斥模式,提示不同发育阶段的基因沉默可能由不同的表观遗传机制主导。

事实上,研究团队早在2021年就已在小鼠模型中明确, Broad H3K4me3的催化酶是SETD1B(注:H3K4甲基化酶共有7种,其中SETD1B是研究关注最少的)。但缺失SETD1B-broad H3K4me3并不是简单地导致基因表达下调,反而在很多关键基因中出现了不恰当的时序性表达上调,如过渡蛋白基因(Tnp)在早期圆形精子细胞中表达上调。这一现象与领域内关于H3K4me3是活化性修饰的认知不符,也难以与精子形成障碍的表型相自洽,因此,一直困扰着研究团队。直至2023年,团队完成了细分发育阶段的基因表达谱和broad/typical H3K4me3分布谱,才揭开了这一谜团:SETD1B-broad H3K4me3在Pol II分配中充当了“大Boss”角色。具体而言,当“大Boss”不在时,Pol II会被更均匀地分配到typical H3K4me3区域,导致这些基因“表达上调”或是在错误时期“开启表达”。例如,过渡蛋白虽然在中晚期精子细胞中才大量产生以替换组蛋白,但其基因座位在早期圆形精子细胞中已经开放,并伴随低水平的typical H3K4me3修饰。当早期圆形精子细胞的broad H3K4me3缺失后,这些应该晚期表达的基因获得了更多的Pol II,导致其在早期圆形精子细胞中反而出现错误的表达上升。有意思的是,随着发育的推进,这些过渡蛋白基因座位在晚期精子细胞中会形成broad H3K4me3,但缺失SETD1B-broad H3K4me3后,它们又无法在真正需要大量表达时维持高表达量。这就完美地解释了2021年时观察到的“分子表型与生物学表型不符”的悖论,也证明了表观遗传调控的复杂与精妙。
1. 建立小鼠精子发生全过程完整表观遗传图谱
为系统解析精子发生过程中的表观遗传特征,研究团队利用此前建立的精子发生同步化技术,结合荧光标记小鼠模型,成功分离并纯化了涵盖有丝分裂、减数分裂以及精子形成阶段的共11种不同发育阶段的生精细胞。通过系统的表观遗传学分析,包括:多种关键组蛋白修饰(H3K27ac、H3K4me1、H3K4me3、H3K27me3、H3K9me2、H3K9me3、H3K36me3)、DNA甲基化以及染色质可及性,研究人员绘制了覆盖小鼠精子发生全过程的表观遗传精细图谱。这一图谱不仅揭示了各类表观遗传修饰在精子发生过程中的动态变化规律,也为理解不同修饰之间的相互作用提供了重要基础。
2. 发现SETD1B介导的H3K4me3宽泛修饰通过调控转录时序性而控制精子细胞发育
在活跃型组蛋白修饰中,H3K4me3和H3K27ac的全局变化显著高于其他组蛋白修饰,且在细线期/偶线期和圆形精子细胞阶段出现两次显著的升高波。第一波,细线期/偶线期阶段的升高与PRDM9激活和DSB热点形成相关,而第二波发生于圆形精子阶段的升高未曾报道。进一步研究发现,圆形精子阶段H3K4me3的升高主要以“broad H3K4me3”(H3K4me3宽泛修饰)形式存在,其修饰富集区宽度通常大于5 kb(平均8.7kb,覆盖30-50个核小体),远超常规的typical H3K4me3修饰(约1-2 kb)。Broad H3K4me3主要修饰精子形成关键基因的启动子/增强子区域,与这些基因在这一阶段的高表达水平密切相关,且在人类精子细胞中也高度保守。
在所有的7类H3K4甲基化酶中,SETD1B的表达谱与Broad H3K4me3出现时期高度吻合,研究人员推测SETD1B很可能是其催化酶。进一步利用条件性基因敲除小鼠模型,研究人员证实了这一点,并发现SETD1B敲除导致雄性不育。有意思的是,SETD1B仅特异性介导broad H3K4me3,其缺失并不影响typical H3K4me3。机制研究揭示,broad H3K4me3修饰能够招募更多的RNA聚合酶II(Pol II)和转录前起始复合物(PIC)成分,从而促进基因的高效表达。然而,当SETD1B缺失时,broad H3K4me3修饰消失,导致Pol II和PIC组分重新分布到常规的typical H3K4me3修饰区域。这种重新分布出现了两个后果:1)原本在某一时期需要高效表达的基因无法维持表达量;2)大量常规表达的基因由于获得了更多转录机器反而提高了表达量,从而影响了精子形成过程的基因表达时序。例如,在SETD1B缺失的小鼠中,一些早期表达的基因在晚期仍然保持较高的表达水平,而一些晚期表达的基因则提前表达。这些精子发生关键基因的严格阶段特异性遭到破坏:在其该表达的阶段无法被充分表达,在其不该表达的阶段又无法被充分抑制。这种“早期基因在晚期无法被充分关闭,晚期基因在早期被提前表达”的现象,最终导致精子形成过程中的基因表达紊乱,影响了精子的正常发育。这是表观领域关于“H3K4me3-基因表达”轴在“新场景”下获得的“新现象”。值得一提的是,SETD1B-broad H3K4me3缺失后,增强子上的H3K4me3完全消失,而启动子区则留下了常规宽度的H3K4me3。说明圆形精子中增强子和启动子区的H3K4me3的催化机制是不同的。
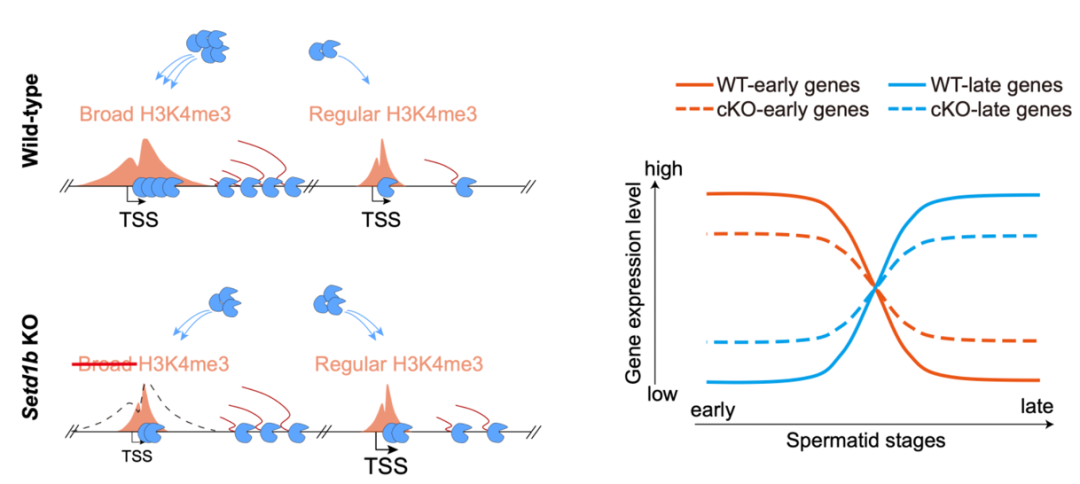
图1:SETD1B介导的Broad H3K4me3调控精子形成中的转录时序性
3. 揭示精子发生阶段特异性的基因沉默机制
该研究进一步揭示了精子发生过程中基因沉默机制的多样性。研究人员观察到,在有丝分裂向减数分裂转换时以及减数分裂重组完成过程中,H3K27me3和H3K9me2这两类异染色质标志发生了两次剧烈的重排。第一次重排始于B型精原细胞,表现为全基因组范围内H3K9me2的显著增加和H3K27me3的全局丢失,而H3K9me3在这些阶段保持相对稳定。分析表明,这种H3K9me2的增加可能与减数分裂基因的沉默有关,尤其是在相对不活跃的基因上更为显著。第二次重排发生在中偶线期粗线期精母细胞(mP),此时减数分裂重组和联会已完成。H3K9me2表现为全局性下调,而H3K27me3水平又迅速重新建立,主要标记那些在mP阶段之前由H3K4me3覆盖、但在mP阶段后需要被沉默的有丝分裂/减数分裂阶段基因。这表明,不同发育阶段的基因沉默由不同的表观遗传机制介导,进一步丰富了对精子发生调控模式的理解。
综上所述,本研究构建了首个覆盖小鼠精子发生全过程的高分辨率表观遗传图谱,全面而系统地揭示了精子发生过程中转录组变化和染色质重塑的表观遗传学基础,为精子生物学研究领域提供了全面完整的表观遗传学基础数据,为生殖生物学和表观遗传学的研究提供了新的研究方向和思路。而SETD1B介导的broad H3K4me3修饰的发现及其功能阐述,为表观遗传修饰如何协调发育过程中基因转录时序性提供了重要见解。
中科院分子细胞科学卓越创新中心林震博士,复旦大学荣博文博士、吕瑞途博士为文章的共同第一作者;中科院分子细胞科学卓越创新中心童明汉研究员,复旦大学生物医学研究院蓝斐教授、北京大学汤富酬教授和复旦大学吕瑞途博士为该文章的共同通讯作者。
原文链接:
https://www.nature.com/articles/s41422-025-01080-0#Abs1