表观团队《Nature》报道METTL3调控小鼠胚胎干细胞异染色质形成机制
mRNA上的m6A修饰受到了广泛的关注,其对于mRNA的命运调控涉及其生物学功能的方方面面,包括剪接、转运、降解、翻译等,m6A主要由METTL3/METTL14复合物催化,目前的研究主要集中在METTL3/METTL14对于细胞质中mRNA的调控研究。但是METTL3是否参与以及如何参与调控染色质功能的研究相对较少。内源性逆转录病毒(Endogenous retrovirus)元件是基因组转座子元件之一,约占小鼠基因组的10%,为了阻止这些转座元件在基因组中四处移动造成遗传突变,细胞进化出了相应的表观遗传修饰抑制这些转座子的活性。小鼠胚胎干细胞主要通过H3K9me3和H4K20me3修饰抑制转座子ERV的活性,但是否还有更多的表观遗传修饰参与其中的研究相对较少。
2021年1月27日,我院沈宏杰青年研究员和牛津大学Yang Shi教授合作在Nature杂志在线发表了METTL3 regulates heterochromatin in mouse embryonic stem cells的研究论文,该研究发现METTL3通过调控内源性逆转录病毒(Endogenous retrovirus)IAPEz亚群上的异染色质状态,进而抑制IAPEz元件转录。

研究发现m6A甲基转移酶METTL3在小鼠胚胎干细胞mESCs染色质主要结合在内源性逆转录病毒(Endogenous retrovirus)IAPEz转座子亚群上。IAPEz转座子主要通过组蛋白H3K9me3和H4K20me3修饰来沉默,H3K9me3主要由甲基转移酶复合物SETDB1/TRIM28复合物催化,H4K40me3则主要由SUV420H1/2催化,并且依赖于H3K9me3。METTL3主要通过招募SETDB1/TRIM28复合物来调控H3K9me3修饰,同时研究者发现METTL3与SETDB1/TRIM28的相互作用不依赖于METTL3的酶活。
同时研究发现METTL3结合IAPEz转座子依赖于METTL3的催化酶活,METTL3的各种酶活突变体都不再结合染色,进一步研究发现METTL3催化的IAPEz RNA促进了m6A识别子蛋白YTHDC1结合染色质,同时YTHDC1反过来促进METTL3在染色质上的结合,形成正向feedback。
研究还讨论了YTH蛋白在裂殖酵母和哺乳细胞中调控异染色质的异同。RNA参与异染色质形成的研究主要集中在裂殖酵母中,裂殖酵母细胞缺少m6A甲基催化酶METTL3同源物,因此YTH同源蛋白Mmi1不识别m6A修饰,而通过识别RNA上的DSR序列参与异染色质形成。而哺乳细胞中YTH同源蛋白YTHDC1通过识别METTL3催化的m6A修饰参与异染色质形成。研究同时还讨论了m6A修饰在RNA的5’UTR和3’UTR可能参与不同的调控作用。
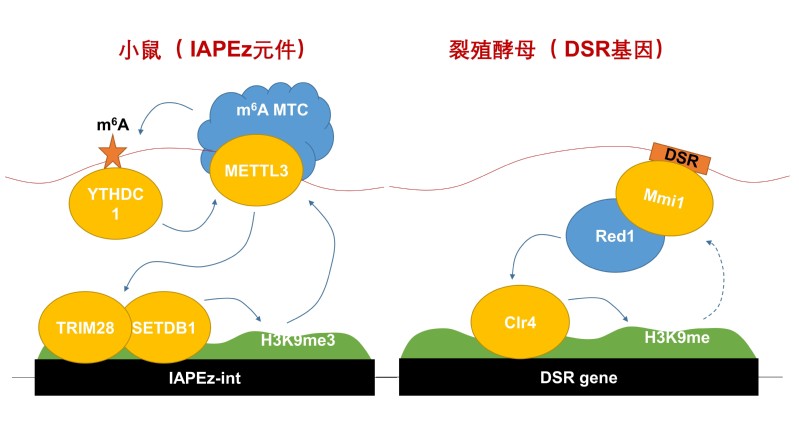
图注:METTL3调控异染色质的工作模型。(左)哺乳细胞中METTL3/YTHDC1蛋白招募H3K9me3甲基转移酶复合物SETDB1/TRIM28的示意图。(右)裂殖酵母细胞中Mmi1蛋白参与调控异染色质形成的示意图。
总之,这项工作发现METTL3可以结合小鼠胚胎干细胞ERV中的IAPEz转座子,通过招募SETDB1/TRIM28维持IAPEz转座子上的异染色质状态。
复旦大学生物医学研究院、上海市医学表观遗传学重点实验室为论文第一署名单位,复旦大学生物医学研究院博士后徐文绮是论文的第一作者,沈宏杰博士和牛津大学Yang Shi教授为论文的共同通讯作者。该项工作受到复旦大学专项资助以及上海市科委,国家自然科学基金委资助。
原文链接:https://www.nature.com/articles/s41586-021-03210-1